Resumo
Objetivou-se avaliar a composição química, fracionamento de carboidratos e compostos nitrogenados, parâmetros de degradação e fermentação ruminal in vitro de genótipos de palma forrageira. O experimento foi conduzido em delineamento inteiramente casualizado, com 4 genótipos de palma forrageira e 4 repetições. Os genótipos de palma forrageira avaliadas foram: Miúda (Nopalea cochenillifera Salm Dyck), IPA Sertânia (Nopalea cochenillifera), Gigante (Opuntia ficus indica Mill), e Orelha de Elefante Mexicana (Opuntia stricta Haw), todas in natura. As amostras foram coletadas aleatoriamente em pontos distintos na área experimental. Plantas inteiras foram coletadas 24 meses após o estabelecimento da cultura de campo. N. cochenillifera Salm Dyck apresentou os maiores teores de matéria seca, proteína insolúvel em detergente ácido, carboidratos não fibrosos, nutrientes digestíveis totais, energia digestível, carboidratos totais e fração A + B1 e menores teores de fibra em detergente neutro corrigida para cinza e proteína, fibra em detergente ácido e celulose (P<0,05). A digestibilidade da fibra em detergente neutro verdadeira in vitro foi elevada para os genótipos N. cochenillifera Salm Dyck e N. cochenillifera Dyck, que também apresentaram altos valores de produção de gás in vitro (P<0,05). Os genótipos de palma forrageira apresentam características químicas adequadas para compor dietas oferecidas aos ruminantes, no entanto, é necessário uma suplementação para aumentar os teores de matéria seca e fibra. Nopalea cochenillifera Salm Dyck apresenta as maiores proporções de nutrientes digestíveis totais, carboidratos não fibrosos, nitrogênio não protéico e alta produção de gases em relação aos demais genótipos analisados aos 24 meses após o estabelecimento do palmal.
Palavras-chave:
Degradabilidade; Digestibilidade; Fermentação in vitro; Nopalea; Opuntia
Abstract
The aim of this study was to evaluate the chemical composition, fractionation of carbohydrates and nitrogen compounds, degradation parameters, and in vitro ruminal fermentation of cactus pear genotypes. The experiment was conducted in a completely randomized design with 4 cactus pear genotypes and 4 replicates. The evaluated cactus pear genotypes were: Miúda (Nopalea cochenillifera Salm Dyck), IPA Sertânia (Nopalea cochenillifera), Gigante (Opuntia ficus indica Mill), and Orelha de Elefante Mexicana (Opuntia stricta Haw), all in natura. Samples were randomly collected at different points in the experimental area. Whole plants were collected 24 months after field crop establishment. N. cochenillifera Salm Dyck presented the highest dry matter, acid detergent insoluble protein, non-fibrous carbohydrate, total digestible nutrients, digestible energy, total carbohydrates, and fractions A + B1 (P<0.05), while presenting lower neutral detergent fiber corrected for ash and protein, acid detergent fiber, and cellulose in relation to the other genotypes studied (P<0.05). The in vitro true digestibility of neutral detergent fiber was high for the genotypes N. cochenillifera Salm Dyck and N. cochenillifera Dyck, which also presented high in vitro total gas production (P<0.05). The cactus pear genotypes show adequate chemical characteristics to be composed part of diets offered to ruminants. However, supplementation is necessary to increase the dry matter and fiber contents. The Nopalea cochenillifera Salm Dyck genotype presented the highest proportions of total digestible nutrients, non-fibrous carbohydrates, non-protein nitrogen, unavailable nitrogen fraction total, and high gas production in relation to the other analyzed genotypes 24 months after field crop establishment.
Keywords:
Degradability; Digestibility; in vitro fermentation; Nopalea; Opuntia
Introdução
O Semiárido brasileiro abrange uma área de aproximadamente 969.589,4 km2, o que corresponde a 11,39% do território nacional(11 Alves FAL, Andrade AP, Bruno RLA, Santos DC, Magalhães ALR, Silva DS. Chemical and nutritional variability of cactus pear cladodes, general Opuntia and Nopalea. Am. J. Food Techn. 2017; 12(1):25-34. Disponível em: https://doi.org/10.3923/ajft.2017.25.34.
https://doi.org/10.3923/ajft.2017.25.34...
). O Nordeste brasileiro possui 70% de sua área inserida no semiárido, caracterizado pela baixa pluviosidade, o que dificulta o estabelecimento de atividades agrícolas(22 Alvalá RCS, Cunha APMA, Brito SSB, Seluchi ME, Marengo JA, Moraes OLL, Carvalho MA. Drought monitoring in the Brazilian Semiarid region. An. Acad. Bras. Ci. 2019; 9(1):e20170209. Disponível em: http://dx.doi.org/10.1590/0001-3765201720170209.
http://dx.doi.org/10.1590/0001-376520172...
). Nessas condições, a vegetação local tende a diminuir ou cessar sua produção de biomassa, ao mesmo tempo em que reduz o valor nutricional, na maioria dos casos. Com isso, há uma irregularidade na oferta de recursos, tornando a produção de alimentos para a pecuária o principal desafio nesta região(33 Magalhães ALR, Sousa DR, Nascimento Júnior JRS, Gois GC, Campos FS, Santos KC, Nascimento DB, Oliveira LP. Intake, digestibility and rumen parameters in sheep fed with common bean residue and cactus pear. Biol. Rhyt. Res. 2019a; 50(1): 1-11. Disponível em: https://doi.org/10.1080/09291016.2019.1592351.
https://doi.org/10.1080/09291016.2019.15...
). Assim, torna-se necessário otimizar recursos para garantir o abastecimento de alimentos, principalmente em tempos de escassez.
A palma forrageira tem sido uma alternativa amplamente utilizada pelos agricultores por ser rica em energia, apresentar adaptabilidade e alto potencial para produção de biomassa no semiárido(44 Louhaichi M, Kumar S, Tiwari S, Islam M, Hassan S, Yadav OP, Dayal D, Moyo H, Dev R, Sarker A. Adoption and utilization of cactus pear in South Asia-Smallholder farmers' perceptions. Sustain. 2018; 10(10):1-15. Disponível em: https://doi.org/10.3390/su10103625.
https://doi.org/10.3390/su10103625...
). Genótipos de palma forrageira pertencentes aos gêneros Opuntia e Nopalea possuem características e propriedades anatômicas e fisiológicas que permitem seu crescimento e desenvolvimento em regiões de terras secas e solos pobres(55 Mendoza PV, Sousa TC, Santos MVF, Mendoza OVV, Dubeux Junior JCB, Lira MA. Organic matter fertilization improves morphological variables in Nopalea cochenillifera Salm Dyck cv. Miúda grown as forage in Pernambuco, Brazil. Rev. Mex. Ci. Pec. 2019; 10(3):756-766. Disponível em: https://doi.org/10.22319/rmcp.v10i3.4386.
https://doi.org/10.22319/rmcp.v10i3.4386...
). Alves et al.(66 Alves FAL, Santos DC, Silva SMS, Silva MC, Mello DR. Metabolismo fotossintético de variedades de palma forrageira cultivadas no Semiárido brasileiro. Pesq. Agropec. Pernamb. 2020; 25(2):e2262252020. Disponível em: https://doi.org/10.12661/pap.2020.009.
https://doi.org/10.12661/pap.2020.009...
) estimam que no território nacional, cerca de 147.439 ha são cultivados com Opuntia fícus-indica (cv. Gigante, Redonda e Clone IPA-20), Opuntia stricta Haw (cv. Orelha de Elefante Mexicana ), Nopalea cochenillifera Dyck (cv. IPA-Sertânia) e Nopalea cochenillifera Salm Dyck (Miúda). Esses genótipos são usados para a produção de frutas e vegetais, consumo humano, ração animal, conservação do solo, biomassa para produção de energia, cochonilha para produção de carmim e como ingrediente na composição de bebidas, medicamentos e cosméticos(11 Alves FAL, Andrade AP, Bruno RLA, Santos DC, Magalhães ALR, Silva DS. Chemical and nutritional variability of cactus pear cladodes, general Opuntia and Nopalea. Am. J. Food Techn. 2017; 12(1):25-34. Disponível em: https://doi.org/10.3923/ajft.2017.25.34.
https://doi.org/10.3923/ajft.2017.25.34...
).
Um dos principais atributos da palma forrageira é o alto teor de água (89%(77 Matias AGS, Araújo GGL, Campos FS, Moraes SA, Gois GG, Silva TS, Emerenciano Neto JV, Voltolini TV. Fermentation profile and nutritional quality of silages composed of cactus pear and maniçoba for goat feeding. J. Agr. Sci. 2020; 158(4):1-9. Disponível em: https://doi.org/10.1017/S0021859620000581
https://doi.org/10.1017/S002185962000058...
)). Assim, esse alimento representa uma reserva estratégica para a conservação da água oferecida aos ruminantes da região semiárida, que sofre constantemente com irregularidades na disponibilidade de água potável para esses animais(11 Alves FAL, Andrade AP, Bruno RLA, Santos DC, Magalhães ALR, Silva DS. Chemical and nutritional variability of cactus pear cladodes, general Opuntia and Nopalea. Am. J. Food Techn. 2017; 12(1):25-34. Disponível em: https://doi.org/10.3923/ajft.2017.25.34.
https://doi.org/10.3923/ajft.2017.25.34...
). Além disso, possui baixos teores de fibra em detergente neutro (304 g/kg de matéria seca) e fibra de detergente ácido (159,7 g/kg de matéria seca)(77 Matias AGS, Araújo GGL, Campos FS, Moraes SA, Gois GG, Silva TS, Emerenciano Neto JV, Voltolini TV. Fermentation profile and nutritional quality of silages composed of cactus pear and maniçoba for goat feeding. J. Agr. Sci. 2020; 158(4):1-9. Disponível em: https://doi.org/10.1017/S0021859620000581
https://doi.org/10.1017/S002185962000058...
) e alto teor de carboidratos não fibrosos (471,5 g/kg de matéria seca(88 Pinho RMA, Santos EM, Oliveira JS, Carvalho GGP, Silva TC, Macêdo AJS, Corrêa YR, Zanine AM. Does the level of forage neutral detergent fiber affect the ruminal fermentation, digestibility and feeding behavior of goats fed cactus pear? An. Sci. J. 2018; 89(10):1-8. Disponível em: https://doi.org/10.1111/asj.13043.
https://doi.org/10.1111/asj.13043...
)). Embora seja um alimento rotineiramente utilizado para alimentação de ruminantes no Semiárido Brasileiro, até onde sabemos, estudos comparativos sobre fracionamento de carboidratos, compostos nitrogenados e parâmetros cinéticos da degradação ruminal in vitro de diferentes genótipos de palma forrageira são escassos(99 Cordova-Torres AV, Mendoza-Mendoza JC, Bernal-Santos G, García-Gasca T, Kawas JR, Costa RG, Jacobo CM, Andrade-Montemayor HM. Nutritional composition, in vitro degradability and gas production of Opuntia ficus indica and four other wild cacti species. Life Sci. J. 2015; 12(2s):42-54. Disponível em: http://dx.doi.org/10.7537/marslsj1202s15.07.
http://dx.doi.org/10.7537/marslsj1202s15...
,1010 Magalhães ALR, Teodoro AL, Gois G.C., Campos, F.S., Souza, J.S.R., Andrade, A.P., Lima, I.E., Oliveira Lp, Nascimento DB. Chemical and mineral composition, kinetics of degradation and in vitro gas production of native cactus. J. Agric. Stud. 2019b; 7(4):129-137. Disponível em: https://doi.org/10.5296/jas.v7i4.15315.
https://doi.org/10.5296/jas.v7i4.15315...
).
A determinação das frações de carboidratos e proteínas e dos parâmetros cinéticos de degradação ruminal é de extrema importância para o nutricionista animal(1111 Santos CB, Costa KAP, Souza WF, Silva VC, Epifanio OS, Santos HS. Protein and carbohydrates fractionation in Paiaguas palisade grass intercropped with grain sorghum in pasture recovery. Acta Scient. An. Sci. 2019; 41(1):1-8. Disponível em: https://doi.org/10.4025/actascianimsci.v41i1.42693.
https://doi.org/10.4025/actascianimsci.v...
). Essas informações podem ser utilizadas em formulações de ração para ruminantes de forma a maximizar a sincronização entre a degradação de carboidratos e compostos nitrogenados, minimizando as perdas de energia e nitrogênio relacionadas a fermentação ruminal e promovendo uma maior eficiência da síntese microbiana(1212 Harun AY, Sali K. Factors affecting rumen microbial protein synthesis: A review. Veterinary Medicine Open J. 2019; 4(1):27-35. Disponível em: https://dx.doi.org/10.17140/VMOJ-4-133.
https://dx.doi.org/10.17140/VMOJ-4-133...
).
Objetivou-se avaliar a composição química, fracionamento de carboidratos e compostos nitrogenados, parâmetros de degradação e fermentação ruminal in vitro de genótipos de palma forrageira disponíveis para ruminantes no Semiárido Brasileiro.
Material e métodos
Análises laboratoriais
As análises laboratoriais foram conduzidas na Universidade Federal Rural de Pernambuco, Unidade Acadêmica de Garanhuns (UFRPE - UAG), em Garanhuns, Pernambuco - Brasil, localizada no Agreste Meridional de Pernambuco (8º 53’ 25’’ latitude sul e 36º 29’ 34’’ longitude oeste, com 896 metros acima do nível do mar). O clima da região é classificado como tropical do tipo Aw', segundo a classificação climática de Köppen e Geiger(1313 Köppen W, Geiger RG. Klimate der Erde. Gotha: Verlag Justus Perthes, Wall-map 150 cm × 200 cm. 1928.), com temperatura média anual de 21,2 ºC e precipitação média anual de 897 mm, sendo os meses Maio, Junho, Julho e Agosto os mais chuvosos. A região é caracterizada por verões quentes e secos e invernos amenos e úmidos, com umidade relativa variando de 75 a 83%. O solo da região é classificado como um Neossolo Regolítico eutrófico típico(1414 Embrapa. Empresa brasileira de pesquisa agropecuária. Sistema brasileiro de classificação de solos, 5th ed. Brasília: Centro Nacional de Pesquisa de Solos; 2018. 356p. Portuguese.).
Amostras e delineamento experimental
O experimento foi conduzido em delineamento experimental inteiramente casualizado, com 4 genótipos de palma forrageira (tratamentos) e 4 repetições por genótipo. Os genótipos de palma forrageira avaliados foram: Nopalea cochenillifera Salm Dyck, Nopalea cochenillifera Dyck, Opuntia ficus indica Mill e Opuntia stricta Haw, todas in natura. As amostras foram coletadas na Estação Experimental de São Bento do Una, Pernambuco - Brasil, pertencente ao Instituto Agronômico de Pernambuco (IPA), localizado na mesorregião do Vale do Ipojuca (8º 31’ 16” latitude sul e 36º 33’ 0” longitude oeste, com 614 metros acima do nível do mar). O clima é classificado, segundo Köppen e Geiger(1313 Köppen W, Geiger RG. Klimate der Erde. Gotha: Verlag Justus Perthes, Wall-map 150 cm × 200 cm. 1928.), como BSwh’ (Tropical Semiárido, quente e seco), com chuvas concentradas no verão, sendo que as maiores precipitações ocorrem entre Janeiro e Março. A estação seca se inicia em Maio e se prolonga até Dezembro e as menores precipitações são verificadas nos meses de Setembro e Outubro. O solo da região é classificado como um Neossolo Regolítico(1414 Embrapa. Empresa brasileira de pesquisa agropecuária. Sistema brasileiro de classificação de solos, 5th ed. Brasília: Centro Nacional de Pesquisa de Solos; 2018. 356p. Portuguese.). A precipitação média no ano das coletas foi de 700,8 mm.
As amostras foram coletadas aleatoriamente de quatro pontos distintos nos campos de cultivo (aproximadamente 0,5 ha) de cada genótipo de palma forrageira, perfazendo quatro repetições por amostra de cada genótipo. As plantas de borda (2 linhas externas) de cada parcela de cultivo e as plantas estabelecidas nas extremidades das linhas intermediárias foram desconsideradas durante a coleta. Plantas inteiras foram coletadas (cerca de 5 cm do nível do solo), 24 meses após o estabelecimento da cultura no campo.
Preparação das amostras para análise
Os genótipos de palma forrageira foram picados com o auxílio de uma faca, homogeneizados manualmente e material picado foi distribuído em bandejas plásticas. As amostras foram pré-secas em estufa de ventilação forçada a 55 °C por 72-h e moídas em partículas de 1 mm e 2 mm (moinho Wiley, Marconi, MA - 580, Piracicaba, Brasil) para determinação da composição química, fracionamento dos carboidratos e compostos nitrogenados e para os ensaios de produção de gás, degradabilidade e digestibilidade in vitro.
Análises químicas
Todas as análises químicas foram realizadas utilizando os procedimentos descritos pela AOAC(1515 Aoac. Association of Official Analytical Chemists. Official methods of analysis, 20th ed. Washington, D.C.: Latimer Jr., G.W.; 2016. 3172p.) para matéria seca (MS, método 967.03), matéria orgânica (MO), matéria mineral (MM, método 942.05), extrato etéreo (EE, método 920.29) e proteína bruta (PB, método 981.10). A fibra em detergente neutro (FDN) e a fibra em detergente ácido (FDA) foram determinadas de acordo com a metodologia proposta por Van Soest et al.(1616 Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polyssacharides in relation to animal nutrition. J. Dairy Sci. 1991; 74(10):3583-3597. Disponível em: https://doi.org/10.3168/jds.S0022-0302(91)78551-2.
https://doi.org/10.3168/jds.S0022-0302(9...
). A fibra em detergente neutro corrigida para cinzas e proteína (FDNcp) foi analisada usando alfa-amilase termoestável, sem sulfito de sódio(1717 Licitra G, Hernandez TM, Van Soest PJ. Standardization of procedures for nitrogen fracionation of ruminant feed. An. Feed Sci. Techn. 1996; 57(4):347-358. Disponível em: https://doi.org/10.1016/0377-8401(95)00837-3.
https://doi.org/10.1016/0377-8401(95)008...
,1818 Mertens DR. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: collaborative study. J. AOAC Int. 2002; 85(6):1217-1240. Disponível em: https://pubmed.ncbi.nlm.nih.gov/12477183/.
https://pubmed.ncbi.nlm.nih.gov/12477183...
) e a lignina foi determinada pelo tratamento do resíduo da fibra em detergente ácido com ácido sulfúrico 72%(1919 Silva DJ, Queiroz AC. Análise de alimentos: métodos químicos e biológicos, 3th ed. Viçosa: Editora UFV; 2006. 235p.). A pectina foi quantificada de acordo com Canteri-Shemin et al.(2020 Canteri-Schemin MH, Fertonani HCR, Waszczynskyj N, Wosiacki G. Extraction of pectin from apple pomace. Braz. Arch. Biol. Techn. 2005; 48(2):259-66. Disponível em: http://dx.doi.org/10.1590/S1516-89132005000200013.
http://dx.doi.org/10.1590/S1516-89132005...
) com alterações de Zanella e Taranto(2121 Zanella K, Taranto OP. Influence of the drying operating conditions on the chemical characteristics of the citric acid extracted pectins from "pera" sweet orange (Citrus sinensis L. Osbeck) albedo and flavedo. J. Food Eng. 2015; 166(1):111-118. Disponível em: https://doi.org/10.1016/j.jfoodeng.2015.05.033.
https://doi.org/10.1016/j.jfoodeng.2015....
). As frações de celulose (CEL) e hemicelulose (HEM) foram estimadas pelas equações: .
Fracionamento dos carboidratos
Os carboidratos totais (CT) foram estimados por meio da equação proposta por Sniffen et al.(2222 Sniffen CJ, O'Connor JD, Van Soest PJ. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992; 70(11):3562-3577. Disponível em: https://doi.org/10.2527/1992.70113562x.
https://doi.org/10.2527/1992.70113562x...
), a saber: . Os carboidratos não fibrosos (CNF), correspondente às frações A + B1, foram determinados pela equação . A fração B2 (fibra digestível) foi obtida pela diferença entre o carboidrato total FDNcp e a fração C (fibra indigestível). A fração C foi obtida pelo FDN indigestível após 288 horas de incubação in situ, conforme descrito por Huhtanen et al.(2323 Huhtanen P, Seppälä A, Ots M, Ahvenjärvi S, Rinne M. In vitro gas production profiles to estimate extent and effective first-order rate of neutral detergent fiber digestion in rumen. J. Anim. Sci. 2008; 86(3):651-659. Disponível em: https://dx.doi.org/10.2527/jas.2007-0246.
https://dx.doi.org/10.2527/jas.2007-0246...
).
Compostos nitrogenados
Os níveis de nitrogênio não protéico (NNP), nitrogênio insolúvel em detergente neutro (NIDN) e ácido (NIDA) foram determinados de acordo com Licitra et al.(1717 Licitra G, Hernandez TM, Van Soest PJ. Standardization of procedures for nitrogen fracionation of ruminant feed. An. Feed Sci. Techn. 1996; 57(4):347-358. Disponível em: https://doi.org/10.1016/0377-8401(95)00837-3.
https://doi.org/10.1016/0377-8401(95)008...
). O fracionamento da proteína foi calculado pelo sistema CNCPS(2222 Sniffen CJ, O'Connor JD, Van Soest PJ. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992; 70(11):3562-3577. Disponível em: https://doi.org/10.2527/1992.70113562x.
https://doi.org/10.2527/1992.70113562x...
). A proteína foi analisada e calculada em cinco frações, A, B1, B2, B3 e C. A fração A (NNP), foi determinada pela diferença entre o N total e o nitrogênio insolúvel em ácido tricloroacético (TCA), com a fórmula: , onde Nt = nitrogênio total da amostra e N1 = teor de nitrogênio insolúvel em ácido tricloroacético.
A fração B1 refere-se à proteína solúvel rapidamente degradada no rúmen, obtida pela diferença entre o nitrogênio insolúvel do tampão borato-fosfato (TBF) menos o NNP, pela fórmula: , onde: N2 = nitrogênio insolúvel em tampão borato-fosfato. As frações B2 e B3 consistem em proteína insolúvel com taxa de degradação intermediária-lenta no rúmen, determinada pela diferença entre o nitrogênio insolúvel do tampão borato-fosfato e o NIDN, o NIDN menos o NIDA, respectivamente. O valor da fração B2 foi determinado pela equação: , e a fração B3: . A fração C é formada pela proteína insolúvel indigestível no rúmen e no intestino e foi determinada pelo teor de nitrogênio residual da amostra após tratamento com detergente ácido e expresso em porcentagem de Nt da amostra.
Os nutrientes digestíveis totais (NDT) foram estimados de acordo com o NRC(2424 Nrc. Nutrient requirements of dairy cattle, 7th ed. Washington, D.C.: National Academy Press; 2001. 381p.) pela equação: , onde 7 é o NDT fecal metabólico; CNFd são carboidratos não fibrosos digestíveis; PBd é a proteína bruta digestível; AGd são ácidos graxos digestíveis; FDNnd corresponde a FDN corrigida para nitrogênio digestível. Para o cálculo de CNFd utilizou-se a equação: , onde PAF é um fator de ajuste igual a 1 para todos os genótipos de acordo com NRC(2424 Nrc. Nutrient requirements of dairy cattle, 7th ed. Washington, D.C.: National Academy Press; 2001. 381p.). Para o cálculo de PBd e AGd foram utilizadas as equações: , levando em consideração que para alimentos com teores de EE < 1, AGd = 0. A FDNd foi obtida em ensaio de degrabilidade in vitro por 48 horas conforme NRC(2424 Nrc. Nutrient requirements of dairy cattle, 7th ed. Washington, D.C.: National Academy Press; 2001. 381p.). A energia digestível (DE) foi estimada pela equação(2424 Nrc. Nutrient requirements of dairy cattle, 7th ed. Washington, D.C.: National Academy Press; 2001. 381p.): .
Cinética da produção de gases e degradabilidade ruminal "in vitro"
A degradabilidade foi determinada de acordo com Tilley e Terry(2525 Tilley JMA, Terry RA. A two-stage technique for the in vitro digestion of forage crops. J. Brit. Grass. Soc. 1963; 18(2):104-111. Disponível em: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2494.1963.tb00335.x.
https://onlinelibrary.wiley.com/doi/abs/...
) através da incubação in vitro de 600 mg de amostra seca ao ar com 60 mL de solução tampão (combinação das soluções A + B com pH 6,8) e 15 mL de inóculo coletado de dois caprinos fistulados no rúmen, filtrado em quatro camadas de gaze, injetando constantemente gás carbônico para manter o ambiente anaeróbico. As incubações ocorreram nos tempos 0, 3, 6, 9, 12, 18, 24, 36 e 48 horas. As amostras no tempo 0 passaram apenas por uma lavagem com água destilada a 39 °C. Os demais tempos foram incubados em estufa com temperatura constante de 39 °C.
Para digestibilidade verdadeira in vitro da matéria seca (DVIVMS), adotou-se a metodologia dos dois estágios descrita por Tilley e Terry(2525 Tilley JMA, Terry RA. A two-stage technique for the in vitro digestion of forage crops. J. Brit. Grass. Soc. 1963; 18(2):104-111. Disponível em: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2494.1963.tb00335.x.
https://onlinelibrary.wiley.com/doi/abs/...
) considerando as modificações propostas por Holden(2626 Holden LA. Comparison of methods of in vitro dry matter digestibility for ten feeds. J. Dairy Sci. 1999; 82(8):1791-1795. Disponível em: https://dx.doi.org/10.3168/jds.S0022-0302(99)75409-3.
https://dx.doi.org/10.3168/jds.S0022-030...
) através de incubações in vitro de 1 g de amostra seca ao ar, com 80 mL de solução tampão e 20 mL de fluido ruminal, filtrados em quatro camadas de gaze, injetando constantemente dióxido de carbono para manter o ambiente anaeróbio. Após 48 horas de incubação, 6 mL de ácido clorídrico (HCl) a 20% e 2 mL de pepsina (1:1000) foram adicionados a cada frasco e, após o período de incubação de 24 horas, foram realizados os procedimentos (filtragem a vácuo, secagem e pesagem dos resíduos) para cálculo do DVIVMS.
Os parâmetros de degradação in vitro (a, b e c) e a degradabilidade potencial da matéria seca foram estimados usando o modelo proposto por Ørskov e McDonald(2727 Ørskov ER, McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agr. Sci. 1979; 92(2):499-503. Disponível em: https://doi.org/10.1017/S0021859600063048.
https://doi.org/10.1017/S002185960006304...
): DP = a + b (1 - e-ct) usando o procedimento PROC NLIN do programa estatístico SAS, onde "PD" é a porcentagem real do nutriente degradado após t horas de incubação, "a" é a fração rapidamente solúvel, "b" é a fração que pode ser degradada se houver tempo, "c" é a velocidade ou taxa de degradação da fração “b” e “t” é o tempo de incubação. Para estimar a degradabilidade efetiva (DE), a seguinte equação foi usada: , onde k é a taxa de passagem do alimento. As taxas de passagem de partículas do rúmen são estimadas em 0,02, 0,05 e 0,08 h-1, conforme sugerido pelo AFRC(2828 Afrc. Agricultural and Food Reaserch Council. Energy and protein requirements of ruminants. Wallingford: CAB International; 1993.159p.).
Para produção de gases in vitro, foi adicionado 1,0 g de amostra em frascos de vidros (160 mL), nos quais foram adicionados 90 mL de meio de cultura conforme Theodorou et al.(2929 Theodorou MK, Williams BA, Dhanoa MS, Mcallan AB, France J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. An. Feed Sci. Techn. 1994; 48(3-4):185-197. Disponível em: https://doi.org/10.1016/0377-8401(94)90171-6.
https://doi.org/10.1016/0377-8401(94)901...
). Na sequência, 10 mL de fluido ruminal (oriundo de três caprinos fistulados no rúmen) foram adicionados em cada frasco, que foram mantidos sob aspersão de CO2. Em seguida os frascos foram vedados com rolhas de borracha e lacres de alumínio. A pressão causada pela fermentação foi aferida usando um transdutor de pressão (Datalogger Universal Logger AG100). As leituras foram feitas em maior frequência durante o período inicial e reduzidas posteriormente (2, 4, 6, 8, 10, 12, 15, 18, 21, 24, 30, 36, 42 e 48 horas de incubação). Os dados de pressão (P) em psi (pressão por polegada quadrada) foram convertidos em volume de gás (V) através da equação , gerada no gerada no Laboratório de Produção de Gases (LPG) da Unidade Acadêmica de Garanhuns - UFRPE, a partir de 937 observações, 1 psi = 4,859 mL de gás. De cada leitura de pressão, foi subtraído o total produzido pelos frascos sem substrato (branco), referentes a cada amostra.
A produção cumulativa de gases foram analisados usando o modelo bicompartimental de Gompertz, citado por Schofield et al.(3030 Schofield P, Pitt RE, Pell AN. Kinetics of fiber digestion from in vitro gas production. Journal of Animal Science. 1994; 72(11):2980-2991. Disponível em: https://doi.org/10.2527/1994.72112980x.
https://doi.org/10.2527/1994.72112980x...
) usando o procedimento NLMIXED do SAS (versão 9.3, SAS Institute Inc., Cary, North Carolina, USA)(3131 Sas. Statistical analysis system institute. SAS/STAT User's guide, version 9.3. Cary: SAS Institute; 2011.): , onde: V(t) = volume máximo total de gases produzidos; Vf1 = volume máximo de gás para a fração de rápida degradação (carboidratos não fibrosos; CNF); Vf2 = volume máximo de gás para a fração de lenta degradação (carboidratos fibrosos; CF); Kd1 = taxa de degradação específica para a fração de rápida degradação; Kd2 = taxa de degradação específica para a fração de lenta degradação; L = duração dos eventos iniciais de digestão (tempo de latência), comum a ambas as fases, e; T = tempo de fermentação (horas).
O modelo bicompartimental de Gompertz foi escolhido assumindo que a taxa de produção de gases é proporcional à atividade microbiana, mas a proporcionalidade diminui com o tempo de incubação, o que pode ser atribuído à perda de eficiência na taxa de fermentação com o tempo(3030 Schofield P, Pitt RE, Pell AN. Kinetics of fiber digestion from in vitro gas production. Journal of Animal Science. 1994; 72(11):2980-2991. Disponível em: https://doi.org/10.2527/1994.72112980x.
https://doi.org/10.2527/1994.72112980x...
).
Análise estatística
Os dados foram submetidos aos testes de Shapiro-Wilk e Levene para verificar a normalidade dos resíduos e homogeneidade das variâncias, respectivamente. Uma vez atendidas as premissas, foram testados por análise de variância utilizando o procedimento General Linear Model (PROC GLM). Os dados da cinética da fermentação in vitro foram analisados usando o procedimento Non-Linear Mixed (PROC NLMIXED), e foram estimados através do método dos mínimos quadrados usando o processo iterativo de Gauss Newton. Os parâmetros de fermentação foram gerados a partir de dados observados em diferentes tempos de incubação in vitro. Todos os dados foram comparados pelo teste de Tukey com nível de significância de 5% usando o software Statistical Analysis System(3131 Sas. Statistical analysis system institute. SAS/STAT User's guide, version 9.3. Cary: SAS Institute; 2011.).
Resultados
A palma forrageira do genótipo N. cochenillifera Salm Dyck apresentou maiores conteúdos de MS (P<0,001), PIDA (P<0,001), CNF (P<0,001), NDT (P= 0,008) e ED (P= 0,003) e menores conteúdos de FDNcp, FDA e CEL (Tabela 1). As for ADF, there were variations from 102.9 g/kg DM for N. cochenillifera Salm Dyck to 188.4 g/kg DM for O. ficus indica Mill (Table 1). MO e pectina foram maiores para os genótipos N. cochenillifera Salm Dyck e O. ficus indica Mill (P<0,05). Os teores de FDNcp foram maiores (P<0,05) para o genótipo O. ficus indica Mill e menores (P<0,05) para o genótipo N. cochenillifera Salm Dyck. A FDA variou de 102,9 g/kg de MS para N. cochenillifera Salm Dyck a 188,4 g/kg de MS para O. ficus indica Mill (Tabela 1). Just as ADL was higher (P<0,05) for the genotype O. ficus indica Mill, although it was low for all genotypes (Table 1). O maior teor de LDA foi observado para o genótipo O. ficus indica Mill (P<0,05; Tabela 1). Para MM, valores mais elevados foram encontrados para os genótipos O. stricta Haw e N. cochenillifera Dyck (P<0,05; Tabela 1). Não foram encontradas diferenças para os conteúdos de EE (P= 0,890; Tabela 1).
A palma forrageira do genótipo N. cochenillifera Salm Dyck apresentou a maior proporção de carboidratos totais (P= 0,0011) e da fração A + B1 (P<0,001) (Table 2). O genótipo O. ficus indica Mill apresentou a maior concentração da fração B2 (fibra potencialmente digestível) (P<0,001). Quanto à fração C, os maiores valores foram observados para o genótipo O. stricta Haw (P<0,001; Tabela 2).
O maior conteúdo de proteína bruta foi observado para os genótipos O. ficus indica Mill e O. stricta Haw em relação a N. cochenillifera Salm Dyck, não diferindo da N. cochenillifera Dyck (P= 0,003). Para o fracionamento dos compostos nitrogenados, o genótipo N. cochenillifera Salm Dyck apresentou as maiores concentrações das frações A (P= 0,001) e C (P<0,001) (Tabela 2). Maiores concentrações da fração B1 + B2, correspondente às frações alta e média de degradação ruminal, foram observadas para os genótipos O. stricta Haw e N. cochenillifera Dyck (P<0,001; Tabela 2). O genótipo O. ficus indica Mill apresentou maior concentração de fração B3 (P <0,001; Tabela 2).
Os maiores valores para fração solúvel de MS in vitro foram observados para o genótipo O. stricta Haw (456,4 g/kg) em relação aos demais genótipos (P <0.001; Tabela 3). A fração lentamente degradável da MS in vitro variou de 423.2 a 640.4 g/kg de MS, com menor valor médio para O. stricta Haw (P <0.001; Tabela 3). O genótipo O. stricta Haw apresentou maior degradabilidade efetiva em função do tempo de incubação de acordo com as taxas lenta, média e rápida (P <0,05; Tabela 3). Não foram observadas diferenças entre os genótipos de palma forrageira para taxa de degradação (c, %/h) (P = 0,070), degradação potencial (P = 0,220) e digestibilidade verdadeira da matéria seca in vitro (P = 0,344) (Tabela 3).
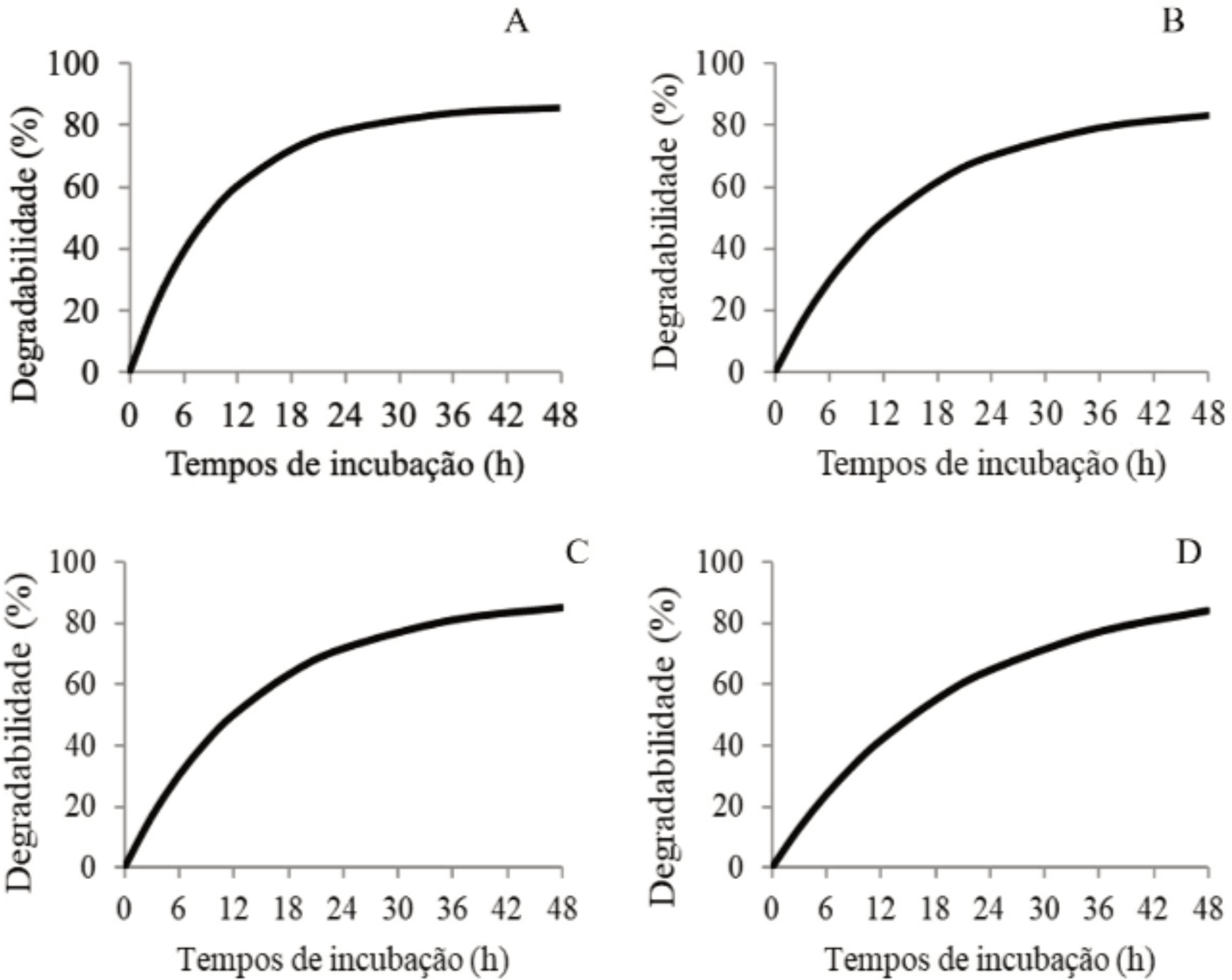
As curvas de degradabilidade em função dos tempos de incubação foram semelhantes para todos os genótipos de palma forrageira (Figura 1).

Curvas de degradabilidade da matéria seca de genótipos de palma forrageira em função dos tempos de incubação in vitro (A - Nopalea cochenillifera Salm Dyck; B - Opuntia ficus indica Mill; C - Opuntia stricta Haw; D - Nopalea cochenillifera Dyck).
O genótipo N. cochenillifera Salm Dyck apresentou o maior volume de gás para a fração de rápida degradação (220,6 mL/g MS; P= 0.008; Tabela 4). Em relação ao tempo de latência (L), os tempos mais elevados foram obtidos pelos genótipos N. cochenillifera Salm Dyck, O. ficus indica Mill e N. cochenillifera Dyck, enquanto o menor tempo foi observado para o genótipo O. stricta Haw (P = 0.001; Tabela 4).
Maiores volumes de gases para a fração de lenta degradação foram observados para os genótipos N. cochenillifera Salm Dyck (115,4 mL/g MS) e N. cochenillifera Dyck (121,7 mL/g MS) (P = 0.001; Tabela 4). O genótipo N. cochenillifera Salm Dyck apresentou a maior produção total de gás (V(t) - 341,1 mL e V(t)2 - 336,0 mL) (Tabela 4). As taxas de degradação Kd1 foram semelhantes para todos os genótipos (P = 0,007; Tabela 4).
As curvas da produção cumulativa de gases dos genótipos de palma forrageira apresentaram forma sigmóide (Figura 2), a qual normalmente é encontrada para as curvas de produção cumulativa de gases em estudos de degradação in vitro. As curvas foram semelhantes entre todos os genótipos estudados. Pode-se observar que, entre 12 e 30 horas de fermentação, as curvas de produção cumulativa de gases apresentaram um decréscimo na produção de gás, evidenciando uma tendência de estabilização após às 30 horas de fermentação.

Produção cumulativa de gases in vitro de genótipos de palma forrageira em função dos tempos de incubação (A - Nopalea cochenillifera Salm Dyck; B - Opuntia ficus indica Mill; C - Opuntia stricta Haw; D - Nopalea cochenillifera Dyck).
Discussão
A palma forrageira apresenta em sua composição baixos teores de matéria seca e proteína e elevados níveis de carboidratos(3232 Al-Arif MA, Suwanti LT, Estoepangestie ATS, Lamid M. The nutrients contents, dry matter digestibility, organic matter digestibility, total digestible nutrient, and NH3 rumen production of three kinds of cattle feeding models. KnE Life Sci. 2017; 36(6):338-343. Disponível em: https://doi.org/10.18502/kls.v3i6.1142.
https://doi.org/10.18502/kls.v3i6.1142...
). Devido às baixas concentrações de MS na palma forrageira, quando há grande oferta desse alimento aos animais, as necessidades de MS podem ser comprometidas. Por outro lado, esta característica representa uma grande contribuição de água para os animais, devido ao alto teor de água que possuem. Isso é benéfico para o Semiárido, que tem a água como fator limitante para a produção animal(3333 Pessoa DV, Andrade AP, Magalhães ALR, Teodoro AL, Santos DC, Araújo GGL, Medeiros AN, Nascimento DB, Valença RL, Cardoso DB. Forage cactus of the genus Opuntia in different with the phenological phase: Nutritional value. J. Arid Env. 2020; 181(104243):1-8. Disponível em: https://doi.org/10.1016/j.jaridenv.2020.104243.
https://doi.org/10.1016/j.jaridenv.2020....
).
A palma forrageira apresenta um alto teor de matéria mineral em sua composição devido a alta concentração de macrominerais, com destaque ao teor de Ca e cristais de oxalato de cálcio(3434 Toit A, Wit M, Hugo A. Cultivar and harvest month influence the nutrient content of Opuntia spp. cactus pear cladode mucilage extracts. Mol. 2018; 23(4):1-12. Disponível em: https://doi.org/10.3390/molecules23040916
https://doi.org/10.3390/molecules2304091...
). Os dados obtidos para MM neste estudo são superiores aos obtidos por Silva et al.(3535 Silva APG, Souza CCE, Ribeiro JES, Santos MCG, Pontes ALS, Madruga MS. Physical, chemical and bromatological characteristics of the giant forage cactus (Opuntia ficus-indica) and small forage cactus (Nopalea cochenillifera) from Paraíba state (Brazil). Rev. Bras. Tec. Agroind. 2015; 9(2):1810-1820. Disponível em: https://doi.org/10.3895/rbta.v9n2.1616
https://doi.org/10.3895/rbta.v9n2.1616...
) que encontraram uma concentração média de MM de 11,9 g/kg MS para o gênero Opuntia e 11,7 g/kg MS para o gênero Nopalea e afirmam que, independente do gênero, a palma forrageira apresenta quantidades consideráveis de MM, porém esses valores variam de acordo com a espécie, idade dos cladódios, área geográfica e época do ano.
Nos genótipos N. cochenillifera Salm Dyck e O. ficus indica Mill mais de 300 g/kg MS é composta por pectina, que representa mais de 50% dos carboidratos não fibrosos, enquanto no genótipo O. stricta Haw esse valor é 25,8 %, ou seja, neste genótipo foi observada apenas metade da pectina presente em N. cochenillifera Salm Dyck e em O. ficus indica Mill. A pectina é um carboidrato estrutural que está presente na parede celular, mas é um carboidrato solúvel em detergente neutro, que faz parte dos carboidratos não fibrosos, portanto de maior potencial de degradabilidade pelos microrganismos ruminais. A alta presença de pectina em genótipos de palma forrageira foi relatada por Morales-Martínez et al.(3636 Morales-Martínez Y, López-Cuellar MR, Chavarría-Hernández N, Rodríguez-Hernández AI. Rheological behaviour of acetylated pectins from cactus pear fruits (Opuntia albicarpa and O. matudae). Food Hydrocol. 2018; 85(1):110-119. Disponível em: https://doi.org/10.1016/j.foodhyd.2018.07.009.
https://doi.org/10.1016/j.foodhyd.2018.0...
).
Os compostos fibrosos da palma forrageira, em geral, são considerados baixos por apresentarem concentração de FDN abaixo de 250 g/kg de MS, característica comum entre os cactos, requerindo a sua associação a uma fonte de fibra de alta efetividade. As diferenças entre os teores de FDA na palma forrageira são explicadas pela quantidade dos carboidratos presentes nessas plantas, que são uma importante fonte de energia para ruminantes que, segundo Van Soest(3737 Van Soest PJ Mertens DR. Nutritional ecology of the ruminant, 2nd ed. Cornell University Press: Ithaca. 1994; 476 p.), os utilizam para o desenvolvimento dos microrganismos que colonizam o rúmen, principalmente carboidratos não fibrosos (açúcar, amido, ácidos orgânicos e frutose), o que a caracteriza como alimento energético, não esquecendo sua adaptação às condições edafoclimáticas da região e alta produção de matéria seca por unidade de área.
Para todos os genótipos testados o conteúdo de NDT foi superior a 650 g/kg MS, confirmando a contribuição energética da palma forrageira para a nutrição animal. Os altos teores de ED e NDT nos diferentes genótipos podem ser explicados pelos altos níveis de carboidratos não fibrosos, que são carboidratos de fácil digestão, como o amido e a pectina.
Os carboidratos totais constituíram mais de 75% da MS. Acima de 90% dos CT apresentaram-se nas frações A e B, indicando que a maioria dos carboidratos está disponível para utilização pelos microrganismos ruminais, confirmando que todos os genótipos estudados podem ser caracterizados como alimentos energéticos. Neste estudo, os genótipos apresentaram baixa concentração da fração C, o que pode resultar em alta digestibilidade dos carboidratos fibrosos(3838 Singh BS, Bhat V, Shukla GP, Singh KK, Gehrana D. Variation in carbohydrate and protein fractions, energy, digestibility and mineral concentrations in stover of sorghum cultivars. Trop. Grass. 2018; 6(1):42-52. Disponível em: https://doi.org/10.17138/TGFT(6)42-52.
https://doi.org/10.17138/TGFT(6)42-52...
). Assim, a importância do fracionamento dos carboidratos ingeridos por ruminantes baseia-se na classificação das bactérias ruminais quanto ao uso dos carboidratos que constituem a parede celular vegetal e daqueles carboidratos localizados no conteúdo celular sem funções estruturais(3939 Deusch S, Camarinha-Silva A, Conrad J, Beifuss U, Rodehutscord M, Seifert J. A structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microb. 2017; 8(1):1-21. Disponível em: https://doi.org/10.3389/fmicb.2017.01605.
https://doi.org/10.3389/fmicb.2017.01605...
-4040 Williams BA, Grant LJ, Gidley MJ, Mikkelsen D. Gut fermentation of dietary fibers: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017; 18(10):1-25. Disponível em: https://doi.org/10.3390/ijms18102203.
https://doi.org/10.3390/ijms18102203...
).
O baixo teor de proteína bruta é uma característica comum em cactáceas. No caso da palma forrageira, isso não é um problema, pois o teor de PB pode ser corrigido associando-o a ingredientes que são fontes de proteína e de nitrogênio não protéico (NNP). Da proteína total, as frações A e B representaram aproximadamente 80,0% nos genótipos O. ficus indica Mill, O. stricta Haw e N. cochenillifera Dyck (Tabela 2). Pode-se afirmar que, embora a palma forrageira se caracterize por apresentar baixa concentração protéica, grande parte dela está disponível para degradação microbiana. A fração C é composta por proteínas relacionadas à lignina, taninos e produtos da reação de Maillard altamente resistentes à degradação microbiana e enzimática, sendo considerada inaproveitável tanto no rúmen quanto no intestino.
Todos os genótipos apresentaram elevados teores de compostos solúveis devido à elevada quantidade de carboidratos de rápida degradação ruminal, em especial os açúcares solúveis e a pectina que essas plantas possuem em sua composição. Para a degradabilidade efetiva estimada de acordo com as taxas de passagem lenta, média e alta, os valores mais altos foram estimados para o genótipo O. stricta Haw. Mesmo com uma taxa de passagem considerada alta, observou-se uma degradabilidade efetiva acima de 55%, o que é satisfatório. Essa degradação potencial é considerada alta porque é uma estimativa da degradabilidade para todos os compostos que podem ser degradados durante o tempo de incubação. A alta digestibilidade verdadeira in vitro da MS de todos os genótipos estudados é um reflexo dos altos teores de carboidratos não fibrosos e baixos teores de lignina, o que pode explicar a elevada taxa de degradação da fração lentamente degradável da MS(4141 Doorenbos J, Martín-Tereso J, Dijkstra J, Van Laar H. Effect of different levels of rapidly degradable carbohydrates calculated by a simple rumen model on performance of lactating dairy cows. J. Dairy Sci. 2017; 100(7):5422-5433. Disponível em: https://doi.org/10.3168/jds.2016-12278.
https://doi.org/10.3168/jds.2016-12278...
).
Todos os genótipos apresentaram DVIVMS de aproximadamente 800 g/kg de MS, sendo considerada satisfatória. Os elevados valores de DVIVMS observados neste estudo estão parcialmente relacionados à quantidade e qualidade dos carboidratos presentes nesses genótipos, uma vez que mais de 75% da MS é composta por carboidratos totais e, destes, mais de 90% são considerados potencialmente digestíveis segundo o fracionamento de carboidratos (Tabela 2). Confirmando o que foi observado neste estudo, Gomez et al.(4242 Gómez LM, Posada SL, Oliveira M. Starch in ruminant diets: a review. Rev. Colomb. Ci. Pec. 2016; 29(2):77-90. Disponível em: https://dx.doi.org/10.17533/udea.rccp.v29n2a01.
https://dx.doi.org/10.17533/udea.rccp.v2...
) relataram que altas proporções de CNF, em geral, aumentam a digestibilidade da palma forrageira. Isso ocorre porque os CNF são prontamente degradados no rúmen, desaparecendo rapidamente e aumentando o suprimento de energia. Isso favorece o crescimento microbiano e, portanto, a digestão.
A alta DVIVFDN dos genótipos N. cochenillifera Salm Dyck e N. cochenillifera Dyck são destacados. Possivelmente o maior resultado seja decorrente do baixo teor de lignina (<3%), que é um constituinte das células vegetais, com digestibilidade baixa ou nula. Portanto, não houve constituintes fibrosos de lenta digestão em quantidade suficiente para dificultar a DVIVMS e DVIVFDN desses genótipos.
O baixo tempo de latência da palma forrageira se deve às suas características físicas e químicas, visto que a fração solúvel é um substrato energético para microrganismos com fermentação rápida, o que facilita os processos de adesão e colonização do substrato. Isso pode aumentar a fermentação dos carboidratos fibrosos e reduzir o tempo de latência(88 Pinho RMA, Santos EM, Oliveira JS, Carvalho GGP, Silva TC, Macêdo AJS, Corrêa YR, Zanine AM. Does the level of forage neutral detergent fiber affect the ruminal fermentation, digestibility and feeding behavior of goats fed cactus pear? An. Sci. J. 2018; 89(10):1-8. Disponível em: https://doi.org/10.1111/asj.13043.
https://doi.org/10.1111/asj.13043...
).
A alta produção de gases observada no genótipo N. cochenillifera Salm Dyck ocorreu porque este genótipo contém altas proporções de CNF e pectina. Os altos volumes de gás dos CF podem ser explicados pela alta DVIVFDN dos genótipos N. cochenillifera Salm Dyck e N. cochenillifera Dyck, uma vez que FDN mais digestível influência a produção de gases a partir dos CF. Portanto, a fibra presente nesses dois genótipos pode ser utilizada por microrganismos ruminais e gerar altas proporções de energia para os animais.
É sabido que a produção de gases in vitro resulta quase que inteiramente dos carboidratos presentes no material incubado e indica degradação por microrganismos ruminais. Assim, N. cochenillifera Salm Dyck apresentou a maior produção total de gases (341,14 mL/g MS) sendo semelhante apenas ao genótipo N. cochenillifera Dyck (303,4 mL/g de MS), ambos pretencentes ao mesmo gênero (Nopalea sp.) (Tabela 4). A maior proporção de carboidratos totais presentes nesses genótipos, o baixo teor de lignina e as baixas frações de carboidratos indisponíveis (fração C) justificam os resultados observados. Além disso, esses mesmos genótipos apresentaram altos valores de DVINFDN, confirmando que a técnica de produção de gases in vitro pode ser utilizada para estimar a qualidade da digestão de carboidratos não fibrosos e fibrosos.
O ajuste do modelo bicompartimental para a geração dos parâmetros de produção de gases in vitro foi considerado o melhor, pois separou a produção dos gases oriundos de carboidratos não fibrosos e fibrosos, gerando uma taxa de degradação para cada um desses componentes. O ajuste desse modelo pode ser observado pela semelhança dos valores reais (V(t)) com os valores estimados pelo modelo (V(t)2). Assim, o modelo bicompartimental é indicado para ajustar a cinética de produção de gases in vitro dos genótipos de palma forrageira estudados.
Conclusão
Os genótipos de palma forrageira apresentam características químicas adequadas para compor dietas oferecidas aos ruminantes, porém, é necessária suplementação para aumentar os teores de matéria seca e fibra. O genótipo Nopalea cochenillifera Salm Dyck apresenta as maiores proporções de nutrientes digestíveis totais, carboidratos não fibrosos, nitrogênio não protéico, fração total de nitrogênio não disponível e alta produção de gases, em relação aos demais genótipos analisados 24 meses após o estabelecimento da cultura no campo.
Agradecimentos
Agradecemos o apoio financeiro do Banco do Nordeste do Brasil (BNB) e a Coordenação de Aperfeiçoamento de Pessoal de Nível Superior no Brasil (CAPES) - Código de Financiamento 001.
References
-
1Alves FAL, Andrade AP, Bruno RLA, Santos DC, Magalhães ALR, Silva DS. Chemical and nutritional variability of cactus pear cladodes, general Opuntia and Nopalea Am. J. Food Techn. 2017; 12(1):25-34. Disponível em: https://doi.org/10.3923/ajft.2017.25.34
» https://doi.org/10.3923/ajft.2017.25.34 -
2Alvalá RCS, Cunha APMA, Brito SSB, Seluchi ME, Marengo JA, Moraes OLL, Carvalho MA. Drought monitoring in the Brazilian Semiarid region. An. Acad. Bras. Ci. 2019; 9(1):e20170209. Disponível em: http://dx.doi.org/10.1590/0001-3765201720170209
» http://dx.doi.org/10.1590/0001-3765201720170209 -
3Magalhães ALR, Sousa DR, Nascimento Júnior JRS, Gois GC, Campos FS, Santos KC, Nascimento DB, Oliveira LP. Intake, digestibility and rumen parameters in sheep fed with common bean residue and cactus pear. Biol. Rhyt. Res. 2019a; 50(1): 1-11. Disponível em: https://doi.org/10.1080/09291016.2019.1592351
» https://doi.org/10.1080/09291016.2019.1592351 -
4Louhaichi M, Kumar S, Tiwari S, Islam M, Hassan S, Yadav OP, Dayal D, Moyo H, Dev R, Sarker A. Adoption and utilization of cactus pear in South Asia-Smallholder farmers' perceptions. Sustain. 2018; 10(10):1-15. Disponível em: https://doi.org/10.3390/su10103625
» https://doi.org/10.3390/su10103625 -
5Mendoza PV, Sousa TC, Santos MVF, Mendoza OVV, Dubeux Junior JCB, Lira MA. Organic matter fertilization improves morphological variables in Nopalea cochenillifera Salm Dyck cv. Miúda grown as forage in Pernambuco, Brazil. Rev. Mex. Ci. Pec. 2019; 10(3):756-766. Disponível em: https://doi.org/10.22319/rmcp.v10i3.4386
» https://doi.org/10.22319/rmcp.v10i3.4386 -
6Alves FAL, Santos DC, Silva SMS, Silva MC, Mello DR. Metabolismo fotossintético de variedades de palma forrageira cultivadas no Semiárido brasileiro. Pesq. Agropec. Pernamb. 2020; 25(2):e2262252020. Disponível em: https://doi.org/10.12661/pap.2020.009
» https://doi.org/10.12661/pap.2020.009 -
7Matias AGS, Araújo GGL, Campos FS, Moraes SA, Gois GG, Silva TS, Emerenciano Neto JV, Voltolini TV. Fermentation profile and nutritional quality of silages composed of cactus pear and maniçoba for goat feeding. J. Agr. Sci. 2020; 158(4):1-9. Disponível em: https://doi.org/10.1017/S0021859620000581
» https://doi.org/10.1017/S0021859620000581 -
8Pinho RMA, Santos EM, Oliveira JS, Carvalho GGP, Silva TC, Macêdo AJS, Corrêa YR, Zanine AM. Does the level of forage neutral detergent fiber affect the ruminal fermentation, digestibility and feeding behavior of goats fed cactus pear? An. Sci. J. 2018; 89(10):1-8. Disponível em: https://doi.org/10.1111/asj.13043
» https://doi.org/10.1111/asj.13043 -
9Cordova-Torres AV, Mendoza-Mendoza JC, Bernal-Santos G, García-Gasca T, Kawas JR, Costa RG, Jacobo CM, Andrade-Montemayor HM. Nutritional composition, in vitro degradability and gas production of Opuntia ficus indica and four other wild cacti species. Life Sci. J. 2015; 12(2s):42-54. Disponível em: http://dx.doi.org/10.7537/marslsj1202s15.07
» http://dx.doi.org/10.7537/marslsj1202s15.07 -
10Magalhães ALR, Teodoro AL, Gois G.C., Campos, F.S., Souza, J.S.R., Andrade, A.P., Lima, I.E., Oliveira Lp, Nascimento DB. Chemical and mineral composition, kinetics of degradation and in vitro gas production of native cactus. J. Agric. Stud. 2019b; 7(4):129-137. Disponível em: https://doi.org/10.5296/jas.v7i4.15315
» https://doi.org/10.5296/jas.v7i4.15315 -
11Santos CB, Costa KAP, Souza WF, Silva VC, Epifanio OS, Santos HS. Protein and carbohydrates fractionation in Paiaguas palisade grass intercropped with grain sorghum in pasture recovery. Acta Scient. An. Sci. 2019; 41(1):1-8. Disponível em: https://doi.org/10.4025/actascianimsci.v41i1.42693
» https://doi.org/10.4025/actascianimsci.v41i1.42693 -
12Harun AY, Sali K. Factors affecting rumen microbial protein synthesis: A review. Veterinary Medicine Open J. 2019; 4(1):27-35. Disponível em: https://dx.doi.org/10.17140/VMOJ-4-133
» https://dx.doi.org/10.17140/VMOJ-4-133 -
13Köppen W, Geiger RG. Klimate der Erde. Gotha: Verlag Justus Perthes, Wall-map 150 cm × 200 cm. 1928.
-
14Embrapa. Empresa brasileira de pesquisa agropecuária. Sistema brasileiro de classificação de solos, 5th ed. Brasília: Centro Nacional de Pesquisa de Solos; 2018. 356p. Portuguese.
-
15Aoac. Association of Official Analytical Chemists. Official methods of analysis, 20th ed. Washington, D.C.: Latimer Jr., G.W.; 2016. 3172p.
-
16Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polyssacharides in relation to animal nutrition. J. Dairy Sci. 1991; 74(10):3583-3597. Disponível em: https://doi.org/10.3168/jds.S0022-0302(91)78551-2
» https://doi.org/10.3168/jds.S0022-0302(91)78551-2 -
17Licitra G, Hernandez TM, Van Soest PJ. Standardization of procedures for nitrogen fracionation of ruminant feed. An. Feed Sci. Techn. 1996; 57(4):347-358. Disponível em: https://doi.org/10.1016/0377-8401(95)00837-3
» https://doi.org/10.1016/0377-8401(95)00837-3 -
18Mertens DR. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: collaborative study. J. AOAC Int. 2002; 85(6):1217-1240. Disponível em: https://pubmed.ncbi.nlm.nih.gov/12477183/
» https://pubmed.ncbi.nlm.nih.gov/12477183/ -
19Silva DJ, Queiroz AC. Análise de alimentos: métodos químicos e biológicos, 3th ed. Viçosa: Editora UFV; 2006. 235p.
-
20Canteri-Schemin MH, Fertonani HCR, Waszczynskyj N, Wosiacki G. Extraction of pectin from apple pomace. Braz. Arch. Biol. Techn. 2005; 48(2):259-66. Disponível em: http://dx.doi.org/10.1590/S1516-89132005000200013
» http://dx.doi.org/10.1590/S1516-89132005000200013 -
21Zanella K, Taranto OP. Influence of the drying operating conditions on the chemical characteristics of the citric acid extracted pectins from "pera" sweet orange (Citrus sinensis L. Osbeck) albedo and flavedo. J. Food Eng. 2015; 166(1):111-118. Disponível em: https://doi.org/10.1016/j.jfoodeng.2015.05.033
» https://doi.org/10.1016/j.jfoodeng.2015.05.033 -
22Sniffen CJ, O'Connor JD, Van Soest PJ. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992; 70(11):3562-3577. Disponível em: https://doi.org/10.2527/1992.70113562x
» https://doi.org/10.2527/1992.70113562x -
23Huhtanen P, Seppälä A, Ots M, Ahvenjärvi S, Rinne M. In vitro gas production profiles to estimate extent and effective first-order rate of neutral detergent fiber digestion in rumen. J. Anim. Sci. 2008; 86(3):651-659. Disponível em: https://dx.doi.org/10.2527/jas.2007-0246
» https://dx.doi.org/10.2527/jas.2007-0246 -
24Nrc. Nutrient requirements of dairy cattle, 7th ed. Washington, D.C.: National Academy Press; 2001. 381p.
-
25Tilley JMA, Terry RA. A two-stage technique for the in vitro digestion of forage crops. J. Brit. Grass. Soc. 1963; 18(2):104-111. Disponível em: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2494.1963.tb00335.x
» https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2494.1963.tb00335.x -
26Holden LA. Comparison of methods of in vitro dry matter digestibility for ten feeds. J. Dairy Sci. 1999; 82(8):1791-1795. Disponível em: https://dx.doi.org/10.3168/jds.S0022-0302(99)75409-3
» https://dx.doi.org/10.3168/jds.S0022-0302(99)75409-3 -
27Ørskov ER, McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agr. Sci. 1979; 92(2):499-503. Disponível em: https://doi.org/10.1017/S0021859600063048
» https://doi.org/10.1017/S0021859600063048 -
28Afrc. Agricultural and Food Reaserch Council. Energy and protein requirements of ruminants. Wallingford: CAB International; 1993.159p.
-
29Theodorou MK, Williams BA, Dhanoa MS, Mcallan AB, France J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. An. Feed Sci. Techn. 1994; 48(3-4):185-197. Disponível em: https://doi.org/10.1016/0377-8401(94)90171-6
» https://doi.org/10.1016/0377-8401(94)90171-6 -
30Schofield P, Pitt RE, Pell AN. Kinetics of fiber digestion from in vitro gas production. Journal of Animal Science. 1994; 72(11):2980-2991. Disponível em: https://doi.org/10.2527/1994.72112980x
» https://doi.org/10.2527/1994.72112980x -
31Sas. Statistical analysis system institute. SAS/STAT User's guide, version 9.3. Cary: SAS Institute; 2011.
-
32Al-Arif MA, Suwanti LT, Estoepangestie ATS, Lamid M. The nutrients contents, dry matter digestibility, organic matter digestibility, total digestible nutrient, and NH3 rumen production of three kinds of cattle feeding models. KnE Life Sci. 2017; 36(6):338-343. Disponível em: https://doi.org/10.18502/kls.v3i6.1142
» https://doi.org/10.18502/kls.v3i6.1142 -
33Pessoa DV, Andrade AP, Magalhães ALR, Teodoro AL, Santos DC, Araújo GGL, Medeiros AN, Nascimento DB, Valença RL, Cardoso DB. Forage cactus of the genus Opuntia in different with the phenological phase: Nutritional value. J. Arid Env. 2020; 181(104243):1-8. Disponível em: https://doi.org/10.1016/j.jaridenv.2020.104243
» https://doi.org/10.1016/j.jaridenv.2020.104243 -
34Toit A, Wit M, Hugo A. Cultivar and harvest month influence the nutrient content of Opuntia spp. cactus pear cladode mucilage extracts. Mol. 2018; 23(4):1-12. Disponível em: https://doi.org/10.3390/molecules23040916
» https://doi.org/10.3390/molecules23040916 -
35Silva APG, Souza CCE, Ribeiro JES, Santos MCG, Pontes ALS, Madruga MS. Physical, chemical and bromatological characteristics of the giant forage cactus (Opuntia ficus-indica) and small forage cactus (Nopalea cochenillifera) from Paraíba state (Brazil). Rev. Bras. Tec. Agroind. 2015; 9(2):1810-1820. Disponível em: https://doi.org/10.3895/rbta.v9n2.1616
» https://doi.org/10.3895/rbta.v9n2.1616 -
36Morales-Martínez Y, López-Cuellar MR, Chavarría-Hernández N, Rodríguez-Hernández AI. Rheological behaviour of acetylated pectins from cactus pear fruits (Opuntia albicarpa and O. matudae). Food Hydrocol. 2018; 85(1):110-119. Disponível em: https://doi.org/10.1016/j.foodhyd.2018.07.009
» https://doi.org/10.1016/j.foodhyd.2018.07.009 -
37Van Soest PJ Mertens DR. Nutritional ecology of the ruminant, 2nd ed. Cornell University Press: Ithaca. 1994; 476 p.
-
38Singh BS, Bhat V, Shukla GP, Singh KK, Gehrana D. Variation in carbohydrate and protein fractions, energy, digestibility and mineral concentrations in stover of sorghum cultivars. Trop. Grass. 2018; 6(1):42-52. Disponível em: https://doi.org/10.17138/TGFT(6)42-52
» https://doi.org/10.17138/TGFT(6)42-52 -
39Deusch S, Camarinha-Silva A, Conrad J, Beifuss U, Rodehutscord M, Seifert J. A structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microb. 2017; 8(1):1-21. Disponível em: https://doi.org/10.3389/fmicb.2017.01605
» https://doi.org/10.3389/fmicb.2017.01605 -
40Williams BA, Grant LJ, Gidley MJ, Mikkelsen D. Gut fermentation of dietary fibers: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017; 18(10):1-25. Disponível em: https://doi.org/10.3390/ijms18102203
» https://doi.org/10.3390/ijms18102203 -
41Doorenbos J, Martín-Tereso J, Dijkstra J, Van Laar H. Effect of different levels of rapidly degradable carbohydrates calculated by a simple rumen model on performance of lactating dairy cows. J. Dairy Sci. 2017; 100(7):5422-5433. Disponível em: https://doi.org/10.3168/jds.2016-12278
» https://doi.org/10.3168/jds.2016-12278 -
42Gómez LM, Posada SL, Oliveira M. Starch in ruminant diets: a review. Rev. Colomb. Ci. Pec. 2016; 29(2):77-90. Disponível em: https://dx.doi.org/10.17533/udea.rccp.v29n2a01
» https://dx.doi.org/10.17533/udea.rccp.v29n2a01
Datas de Publicação
-
Publicação nesta coleção
25 Out 2021 -
Data do Fascículo
2021
Histórico
-
Recebido
16 Jun 2021 -
Aceito
16 Ago 2021 -
Aceito
20 Set 2021